The COVID-19 pandemic has undoubtedly had a profound impact on the world. Its prevalence has been a daily talking point ever since the virus was declared a pandemic in March of 2020 by the World Health Organization. Through discussion around the origins and mechanisms of the disease, the public was exposed to and educated on the basic science behind viruses while simultaneously being reminded of the horrible consequences viruses can have on their host. As COVID-19 is frequently compared to the Influenza A Virus, also more commonly known as the flu, people were also forced to learn about the flu, a disease that many often neglect or fail to acknowledge as anything more than a seasonal illness.
In the past century, there have been a total of four influenza virus pandemics: H1N1 (1918), H2N2 (1957-8), H3N2 (1968), and H1N1 (2009). Collectively, these pandemics have taken tens of millions of lives across the globe. According to the Centers for Disease Control and Prevention (CDC), the number of deaths due to influenza in the U.S. alone is estimated to be around 30,000 annually, as is observed in Figure 1. Globally, seasonal influenza viruses infect anywhere between five and twenty percent of the human population per year. To mitigate and prevent influenza infections, the primary preemptive treatments are vaccinations. However, influenza has the propensity to unpredictably mutate and rearrange itself to create different strains. Consequentially, it is difficult for pharmaceutical companies to correctly anticipate which strains will appear in the next flu season. Therefore, they cannot always manufacture and distribute the most effective vaccines each year [1].
Figure 1

CDC Statistics on the Estimated U.S. Influenza Burden by Season from 2010 to 2020 [1]
In addition to creating preventative measures, researchers are currently looking to develop antiviral treatments that specifically target the influenza virus once it has entered a person’s system. For the drugs to have specificity to the influenza virus, they must target a key component of the virus’ machinery. Fortunately, this component has been identified: the influenza virus RNA-Dependent RNA-Polymerase (RdRp).
Viruses come in many different shapes and sizes. At their core, viruses are infectious segments of nucleic acids, either DNA or RNA, surrounded by a protein and sugar coat that allows them to bind and infect host cells. However, the viruses need to make use of a host’s cell machinery and resources to replicate. Influenza contains its genetic code in the form of a negative sense single strand of RNA (ssRNA). Negative ssRNA cannot be read by the machinery in the cell, so the influenza virus must also have an RdRp with it during infection to generate a positive ssRNA. The positive ssRNA can then be utilized by the cell to make the necessary components to replicate and spread the virus, as depicted in Figure 2 [2]. Hence, the RdRp is a crucial component of the influenza virus life cycle, making it a target for drug therapy.
Figure 2
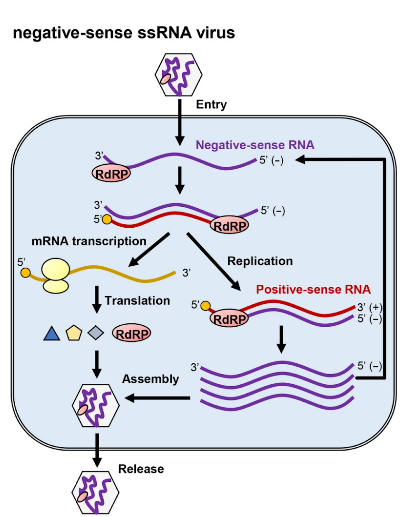
Negative Sense ssRNA Virus Replication Process [2]
The influenza RdRp is comprised of three domains: PA, PB1, and PB2, (Figure 3). The PA domain has endonuclease, or cutting, activity, allowing it to bind and cleave host mRNA produced in the nucleus. This process, known as “cap-snatching,” is performed to create a capped primer, or starting sequence. This primer can then be used to initiate transcription, or the copying of the negative ssRNA into positive ssRNA. This copying step occurs in the PB1 domain. The PB2 domain holds the primer in place for copying to start. Without the cleavage activity of PA, no viral replication could transpire. Therefore, if scientists can produce a drug that blocks this activity by targeting this domain, they can prevent the spread of influenza post-infection [3].
Figure 3
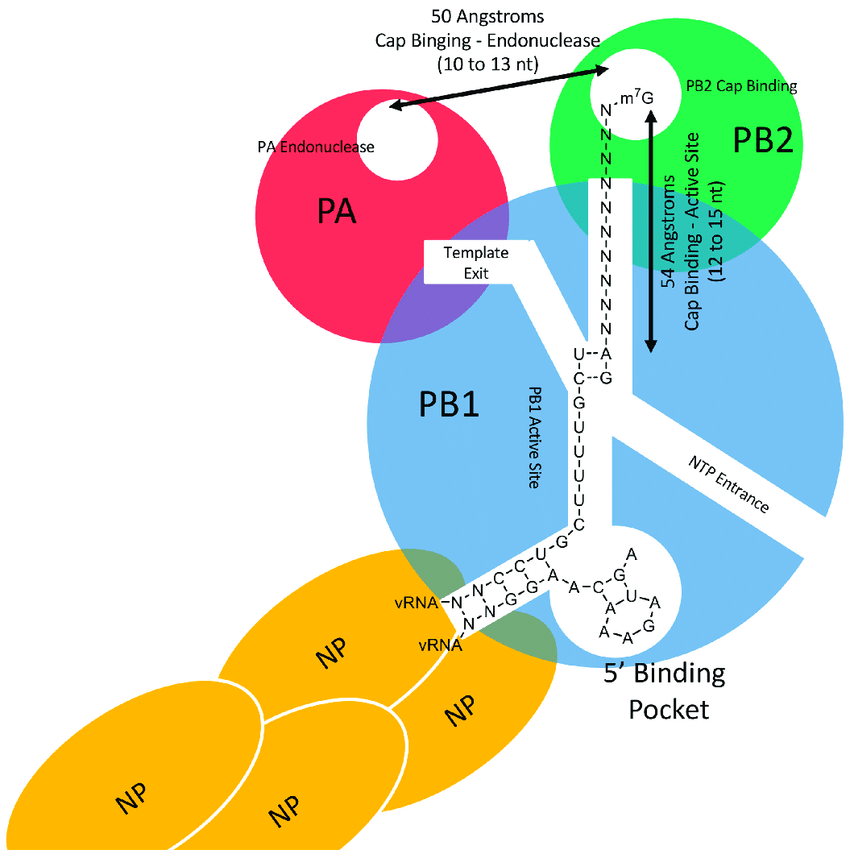
Three Domains of the Influenza RNA-Dependent RNA-Polymerase [3]
In their 1999 paper published in the journal Biochemistry, researchers at Roche DiscoVery Welwyn in the UK outlined the role of divalent metal ions, which have a positive 2 charge, and their importance in the process of RNA cleavage by the influenza RdRp. Metal ions can be critical to the structure and function of a protein as they can stabilize charges and catalyze reactions. Hence, the researchers were interested in which metals most directly impacted the function of the RdRp. This study was performed using influenza RdRp and a known RNA sequence, called Gem20 RNA, as a substrate for the polymerase to make a start sequence primer. The RdRp consistently cleaved Gem20 after the eleventh nucleotide, resulting in a G11 product that the researchers could measure and quantify. In one experiment, they individually added 16 different metal ions with the RNA sequence and RdRp and then looked at the amount of G11 produced. The data collected, found in Table 1, determined that six divalent metal ions (Mn2+, Co2+, Mg2+, Zn2+, Ni2+, and Fe2+) showed significant activation of RNA cleavage. However, only Mn2+ and Mg2+ showed significant activation of the PB1 transcription, or RNA copying, activity [4].
Table 1
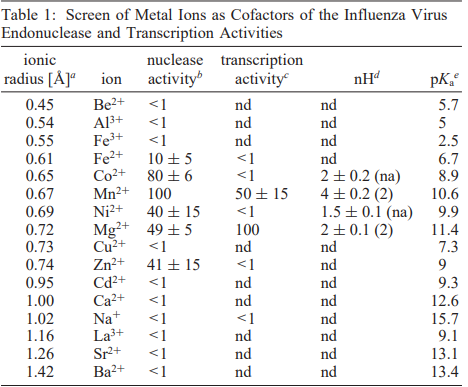
A Screen of Metal Ions as Cofactors of the Influenza Virus Endonuclease and Transcription Activities [4]
The metals which activated RNA cleavage did so in a positively cooperative fashion (nHd greater than 1). This means that when one ion of metal binds to the RdRp, it makes the RdRp more likely to bind another metal ion. This finding also told the researchers that there were at least two metal-binding sites in the RdRp. In this experiment, the metals Mn2+, Co2+, Mg2+, and Ni2+ were found to bind cooperatively to the RdRp for RNA cleavage activation while only Mn2+ snd Mg2+ bound cooperatively for RNA transcription activation (Table 1) [4].
With these discoveries, the researchers wanted to know what would happen to the RNA cleavage activity if they combined different metals together. In Figure 4a, they found that low concentrations of Mn2+ enhanced RNA cleavage in the presence of increasing Mg2+ (white circles) when compared to the cleavage activity with Mg2+ by itself. Figure 4b shows that Mg2+ has a greater activating effect in the presence of a fixed amount of Ni2+ (white circles) or Co2+ (black triangles) than by itself (black circles). Similarly, in Figure 4c, a fixed amount of Mg2+ in a sample with increasing Ni2+ (black circles) led to an increase in cleavage activity compared to a sample with only Ni2+ present (white circles). The researchers propose that the two different metal ions in each case could have different functions. Hence, for optimal cleavage activity, a combination of metals would be required to contribute to their specialized function. In all, these findings provide evidence that combining different metals together can make the influenza RdRp better at cleaving RNA, and thereby better at replicating and spreading itself throughout a host [4].
Figure 4
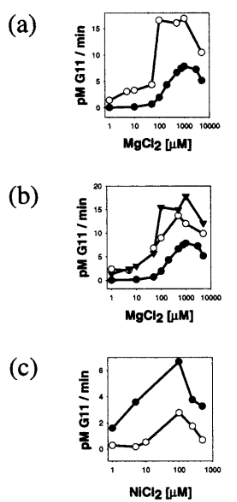
Synergistic Activation of RNA Cleavage in the Presence of Combinations of Different Metal Ions [4]
The overall findings from the 1999 paper in Biochemistry helped piece together the mechanism behind RdRp cleavage activity. As of today, crystal structures of the protein have confirmed the presence of two Mn2+ ions in the active site in the PA domain of RdRp which are vital for RNA cleavage activity to perform the “cap-snatching” mechanism [5]. Additionally, researchers and companies have already begun developing drugs that specifically target the PA domain. One such drug known as L-742,001 is a PA endonuclease inhibitor that binds to and blocks the RNA cleavage active site. This prevents RNA cleavage of host mRNA, thereby stopping the production and spread of the influenza virus. While this drug has recently been deemed unsuitable for further drug development, a research group in the U.S. sought to identify and characterize influenza variants resistant to these types of inhibitors. By doing so, they hoped to learn how to combat future resistant viruses if they ever appeared [5].
The team first induced mutations into wild-type (WT) influenza viruses to create a library of different genetic lines they could evaluate. The viruses were then grown in standard MDCK cells, kidney cells from a female cocker spaniel. Resistant viruses were sequenced, and the four mutations they found in the active site of the cleavage domain were T20A, I79L, E119D, and T20A-F105S [5].
Their next approach was to determine the functional impact the resistance mutations had on the viruses. Therefore, they wanted to look at how the mutations affected the ability of the RdRp to cleave host RNA. To do this, they conducted a TCID50 Assay to determine the number of infectious virus particles that were created in the absence or presence of L-742,001. The assay works by varying the amount of virus added to each group of cells via a serial dilution. In this experiment, viruses that are not resistant to the L-742,001 drug would not be capable of proper replication, thereby resulting in low viral concentrations. This can be observed in the WT influenza virus as it is exposed to the inhibitor (red line) in Figure 5a. In Figure 5 b-d, the T20A, I79L, and F105S, respectively, demonstrate they were partially resistant to the L-742,001 drug. Furthermore, in Figure 5 e-f, the E119D and T20A-F105S were completely unaffected by the inhibitor [5].
Figure 5

Impact of Endonuclease Resistance Mutations on Growth of Influenza Virus [5]
The researchers then added the unaffected mutations from the previous experiment, E119D and F105S, separately to the 2009 H1N1 influenza virus, which already has the T20A mutation. With these new strains, they ran experiments to determine the IC50 for each mutant, or the amount of drug required to inhibit half of the influenza replication. A larger IC50 means more drug is required to combat the virus, indicating the drug is less effective against that particular strain. Their data showed a 4.2-fold increase for the E119D mutation and a 9.7-fold increase for the F105S mutation in the IC50 values for L-742,001, thereby demonstrating that these mutations had made the virus more resistant to the drug. However, as seen in Figure 6a, the RNA cleavage functionality of the E119D mutant (blue line) was completely done away with when compared to the WT virus (pink line). F105S also partially lost its cleavage function (green line). The inability of the aspartic acid (D) in the E119D mutation to properly coordinate one of the two crucial Mn2+ ions in the active site likely contributes to the significant loss of function in the E119D mutant. As previously mentioned, metals play an important role in catalyzing reactions in numerous proteins, such as the influenza RdRp. Compared to a WT active site, if the metal is not properly positioned, it will not be able to function properly.
Figure 6
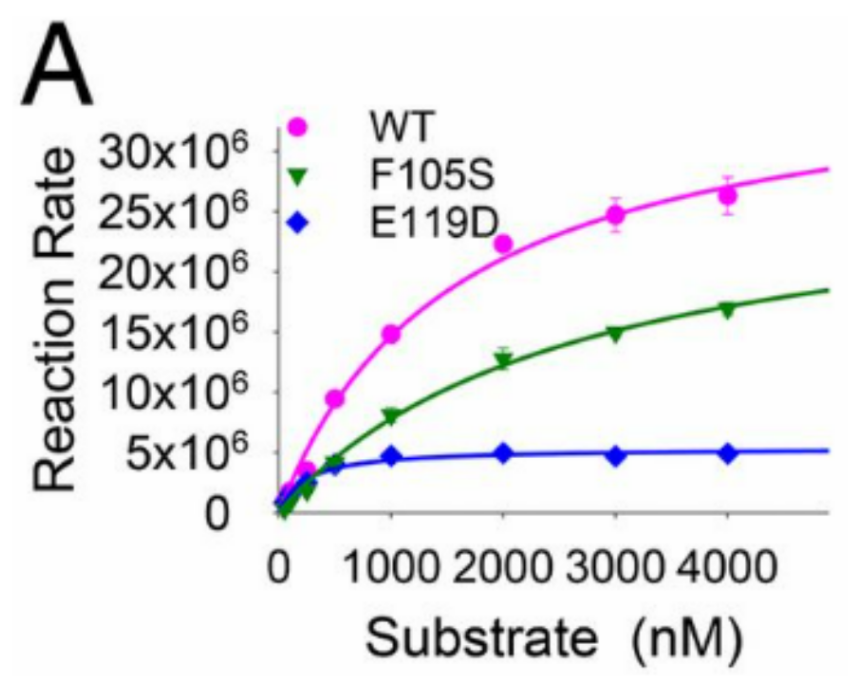
Kinetic Study of the RNA Cleavage Rate by the Influenza RdRp Endonuclease Domain [5]
Given the importance of RNA cleavage activity in influenza replication, a decrease in this function would hinder the competitiveness of mutants compared to other influenza strains. Even the slightest difference could lead to a significant reduction in competitiveness, making both of these mutations unlikely. Additionally, the researchers concede that the specific amino acids they altered in these two instances are highly conserved, or unchanged between different influenza generations and strains. In fact, the two residues they mutated (F105 and E119) are found in more than 99.5% of all human and avian influenza viruses [5]. Therefore, it is unlikely these ‘super’ viruses with resistance to endonuclease inhibitors are going to appear any time soon. Still, this paper provides useful information regarding how this type of resistance could emerge which can hopefully guide future drug discovery and development efforts to design compounds less prone to developing drug resistance.
This article was written by Aidan Jones (contact info: asjones@wesleyan.edu).
Sources:
- “Disease Burden of Flu.” Centers for Disease Control and Prevention, Centers for Disease Control and Prevention, 7 Jan. 2022, https://www.cdc.gov/flu/about/burden/index.html.
- RNA-Dependent RNA Polymerase, RdRP, a Promising … – Wiley Online Library. https://onlinelibrary.wiley.com/doi/10.1111/cas.14618.
- De Vlugt C, Sikora D, Pelchat M. Insight into Influenza: A Virus Cap-Snatching. Viruses. 2018; 10(11):641. https://doi.org/10.3390/v10110641
- Doan, Linh, et al. “Metal Ion Catalysis of RNA Cleavage by the Influenza Virus Endonuclease.” Biochemistry, vol. 38, no. 17, 1999, pp. 5612–5619., https://doi.org/10.1021/bi9828932.
- Song, Min-Suk, et al. “Identification and Characterization of Influenza Variants Resistant to a Viral Endonuclease Inhibitor.” Proceedings of the National Academy of Sciences, vol. 113, no. 13, 2016, pp. 3669–3674., https://doi.org/10.1073/pnas.1519772113.